MicroRNAs in the development of left ventricular remodeling and postmyocardial infarction heart failure
Key words: acute myocardial infarction, circulating microRNAs, heart failure, left ventricular remodeling, prognostic biomarkers



 CC BY-NC-SA 4.0
CC BY-NC-SA 4.0
MicroRNAs in the development of left ventricular remodeling and postmyocardial infarction heart failure
Acute myocardial infarction (AMI) induces unfavorable left ventricular remodeling (LVR), a complex process that involves molecular, cellular, and geometric alterations leading to important changes in heart structure and function. Heart failure (HF) is a frequent complication of AMI, and it remains a serious clinical, epidemiological, and economic challenge. Despite advances in the therapy and management of HF, many patients still suffer from severe symptoms. The underlying molecular mechanisms of the post‑AMI LVR are not yet fully understood. Numerous studies have indicated that dysregulation in the expression of microRNA (miRNA) molecules leads to changes in several pathological processes in the heart, which are associated with post‑AMI transition from cardiac hypertrophy to HF. In this review, we summarize the current knowledge on the role of miRNAs in the regulation of basic processes, such as excessive myocardial fibrosis, pathological cardiomyocyte hypertrophy, and myocardial cell apoptosis. Moreover, the significance of circulating miRNAs as noninvasive prognostic biomarkers in the prediction of LVR and HF after AMI has also been discussed. In conclusion, miR‑29 family members (miR‑29a and miR‑29b), miR‑150, and miR‑30a‑5p represent different groups of miRNAs, but all of them are involved in the regulation of the fundamental processes associated with post‑AMI left ventricular dysfunction and HF. Furthermore, these miRNA molecules may serve as a potential therapeutic target during disease progression.
Introduction
Acute myocardial infarction (AMI) is a life‑threatening condition that frequently leads to heart failure (HF).1,2 It is also associated with unfavorable left ventricular (LV) remodeling (LVR), which affects cardiac function and increases the risk of HF.3 An example of unfavorable LVR after AMI is bendopnea, which is connected with changes in ventricular filling and is one of the symptoms of chronic HF.4 The molecular processes involved in the pathogenesis of HF following AMI include extensive myocardial damage, recurrent ischemia and impairment of myocardial contractility, chronic activation of the neuroendocrine system, and extracellular matrix (ECM) remodeling.1,5 According to the literature, one‑third of patients with ST‑segment elevation MI develop LVR.6 Left ventricular systolic dysfunction and other adverse events often worsen with consecutive stages of HF. For many years, the measurement of natriuretic peptides (especially N‑terminal pro‑B‑type natriuretic peptide [NT‑proBNP]), has been widely used in clinical practice to assess LV systolic dysfunction and monitor treatment effectiveness in patients with HF.7 Gerber et al8 reported that 70% of patients who develop HF after AMI die within 7.6 years. Heart failure remains the leading diagnosis on admission in patients over 65 years old.9 Based on the Framingham Heart Study, a 5‑year mortality rate is 45% to 60%.10,11 Early prediction of adverse LVR and the occurrence of HF after AMI is essential to improve morbidity and mortality outcomes. Despite advances in the identification of biomarkers associated with LVR, the ideal biomarker for an early and reliable prediction of post‑AMI HF is still to be found.12
MicroRNAs (miRNAs, miRs) belong to the best characterized class of noncoding RNAs. For several years, a growing interest in the biology of miRNAs in cardiovascular research has been observed. Some miRNAs are highly expressed in the heart, and numerous studies investigated the significance of miRNAs in cardiac development, physiology, and disease.13-15 An increasing body of evidence indicates that the upregulation and downregulation of miRNAs can be considered a marker of environmental changes in heart disease and HF due to the causative and protective role of these molecules, so important for modulating metabolic pathways.16 The development of new therapies has become possible owing to the engagement of the scientific community in a thorough research of miRNA molecules as a potential therapeutic target, in an attempt to explain the pathophysiology of cardiovascular disease.17 Currently, a considerable interest in circulating miRNA molecules has been observed due to their potential role as diagnostic and prognostic biomarkers of various pathological processes occurring in the heart, including the development of LVR and post‑AMI HF.
MicroRNAs: origin, structure, function, and regulation
MicroRNAs belong to a class of small (~22‑nucleotide long), single‑stranded, and evolutionarily conserved molecules. These noncoding RNAs act as posttranscriptional gene regulators by the inhibition of target mRNA translation or the promotion of mRNA degradation.17-19 Due to broad target interactions, miRNAs participate in the regulation of a wide range of biological processes, such as programmed cell death, metabolism, immune responses, cell proliferation, differentiation, organogenesis, and many others.19-21
According to the miRBase Sequence Database (version 22.1), more than 2000 known human miRNAs are documented.22 It has been estimated that more than 60% of human protein‑coding genes are targeted by a single miRNA.23
The miRNA biogenesis pathway consists of complex processes occurring in the nucleus and cytoplasm (Figure 1). The first process occurs in the nucleus and leads to the generation of long primary miRNAs (pri‑miRNAs), which are generally transcribed by RNA polymerase II.24-27 MicroRNA genes can be expressed as independent transcripts (intergenic miRNA), polycistronic transcripts that often encode multiple miRNAs, or they can be embedded in introns of protein‑encoding mRNAs (intragenic miRNA).28 Pri‑miRNAs consist of hundreds to thousands of nucleotides, and they contain an active miRNA in a stem‑loop structure. Pri‑miRNAs molecules are processed in the nucleus by a microprocessor complex consisting of ribonuclease III (Drosha) and DiGeorge Critical Region 8 (DGCR8) to form a 60- to 100‑nucleotide long hairpin‑shaped precursor miRNAs (pre‑miRNAs).29,30 Pre‑miRNAs are transferred from the nucleus into the cytoplasm by Exportin 5.30,31 In the cytoplasm, pre‑miRNAs are processed by ribonuclease III type endonuclease (Dicer) to small double‑stranded duplex containing the mature miRNA (18- to 25‑nucleotide long).30 Functional miRNAs are finally coupled to Argonaute 2 protein (Ago2) and then incorporated into the RNA‑induced silencing complex (RISC).27 The mechanism of mRNA regulation by miRNA depends on the degree of sequence complementarity between the miRNA and 3’UTR motif in the target mRNA gene.32 The extent of base pairing determines their mode of action: translation inhibition and/or augmented mRNA degradation, both pathways resulting in the effective downregulation of the target gene expression. Some miRNAs may also promote mRNA translation.17,33 Of note, an individual miRNA can regulate the expression of multiple target genes, at the same time being controlled by numerous factors and molecular mechanisms itself, thus creating a complex system of mutual interactions.34-36
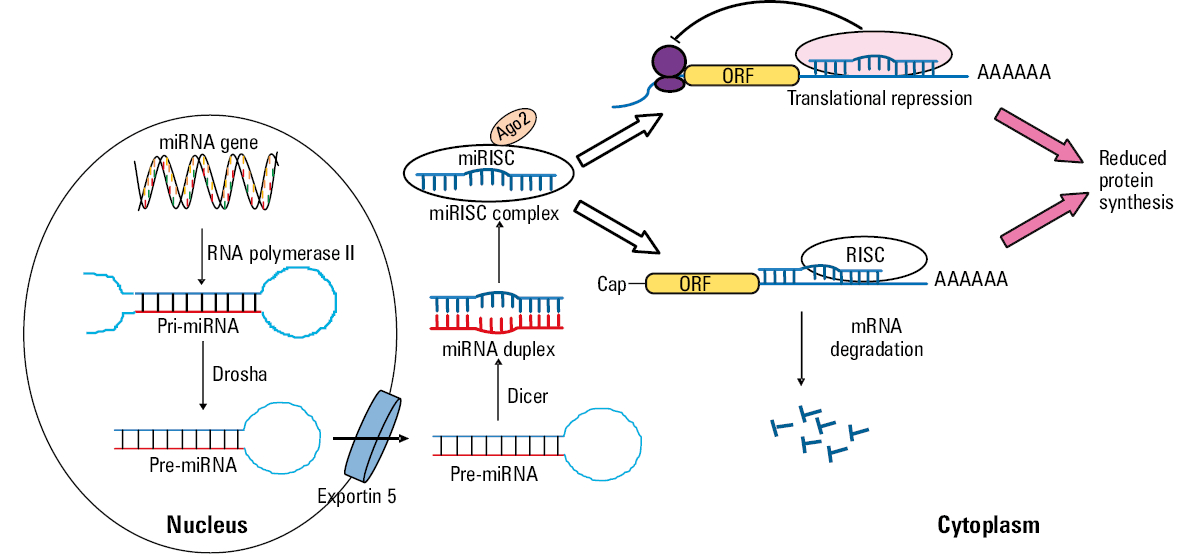
In order to fully elucidate the pathogenesis and treatment of cardiovascular diseases caused by dysregulated gene expression, it is essential to understand the complexity of the mechanisms involved in the regulation of miRNA expression.
MicroRNAs and myocardial infarction
Myocardial infarction is an effect of myocardial ischemia, resulting in myocardial necrosis. Due to the lack of oxygen, the reduced blood flow to the heart may lead to the damage of cardiomyocytes. The above processes are usually caused by the rupture of the atherosclerotic plaque in the coronary vessels.37
The role of tissue‑specific miRNAs both in physiological and pathological conditions has been documented. MicroRNAs are critical regulators that are involved in almost all aspects of cardiovascular diseases. During MI, numerous miRNA molecules are released from cardiomyocytes into the bloodstream.38,39 Cellular miRNAs can be transported into circulation as microvesicles, exosomes, and apoptotic bodies or as miRNA associated with proteins, creating miRNA‑protein complexes with Ago2 or nucleoplasmin, as well as HDL‑miRNA complexes.23 Circulating miR‑1, miR‑133a, miR‑133b, miR‑208a, miR‑208b, and miR‑499 are frequently associated with MI; miR‑133a, miR‑133b, and miR‑1 are strongly expressed in the heart and skeletal muscle, whereas miR‑499 and miR‑208a are only cardiac specific.27,38,40,41
One of the most upregulated miRNAs is miR‑1, while miR‑133a is downregulated in rat hearts upon acute myocardial ischemia–reperfusion.42 Yin et al43 reported that after ischemic preconditioning, miR‑1, miR‑21, and miR‑24 expression was significantly elevated in the heart. The increased miRNA levels trigger cardiac protection by upregulating not only endothelial nitric oxide synthase but also heat‑shock protein 70 and heat‑shock transcription factor 1.43
In our previous study,44 we demonstrated that miR‑208a molecules are released into the circulation (a 3‑fold increase vs control) during AMI, even before the appearance of traditional biomarkers of cardiomyocyte necrosis: cardiac troponin I and creatine kinase‑MBmass. The highest expression of miR‑208a (a 90‑fold increase vs control) was observed in the third hour after reperfusion. Plasma expression of miRNA‑208a correlated with cardiac troponin I and creatine kinase‑MBmass concentrations. These results clearly indicate that miRNA‑208a molecules, which were present in plasma, had been released from cardiac myocytes. After 24 hours, their expression decreased to the level observed on admission.44 This is contrast to the study presented by Zile et al,45 who observed a 3‑fold increase in miRNA‑208 expression as compared with controls only until day 5 after AMI. The maximum expression occurred on day 28, and the increased expression was maintained for the next 90 days.45 This late plasma expression of miR‑208a was associated with the development of HF in these patients. These studies seem to suggest that there are at least 2 different mechanisms of miR‑208 molecule release from cardiomyocytes. In the early phase of MI, miR‑208a may be released in microvesicles formed during myocardial ischemia. The late release is probably associated with cardiomyocyte damage or apoptosis (or both).
MicroRNAs and heart failure
Cardiac remodeling is an adaptive myocardial change that maintains the hemodynamic balance under the influence of external stimuli. In the continuous presence of this causative factor, such remodeling becomes an irreversible process leading to HF.30 The pathological ventricular remodeling is characterized by 3 main features: extensive fibrosis, pathological cardiomyocyte hypertrophy, and myocardial cell apoptosis (Table 1).17 MicroRNA molecules involved in the above processes with a reverse direction of the mechanism of action (anti- and pro-) are shown in Table 1.
Fibrosis | Hypertrophy | Apoptosis | |||
Antifibrotic | Profibrotic | Antihypertrophic | Prohypertrophic | Antiapoptotic | Proapoptotic |
miR‑24 | miR‑21 | miR‑1 | miR‑23a | miR‑30 | miR‑101a |
miR‑29 | miR‑125b | miR‑9 | miR‑27b | miR‑199 | miR‑200a |
miR‑30 | miR‑26 | miR‑199b | miR‑206 | miR‑320 | |
miR‑31 | miR‑133 | miR‑330 | |||
miR‑133 | |||||
Excessive myocardial fibrosis
The balance between collagen synthesis and degradation is essential for myocardial ECM homeostasis. Activated fibroblasts synthetize collagen, which is then degraded by matrix metalloproteinases.17 Several studies documented that miRNA molecules regulate collagen metabolic pathways. Major antifibrotic miRNAs include miR‑24, miR‑29, miR‑30, miR‑31, and miR‑13330 (Table 1), while miR‑133 and miR‑30 regulate the expression of connective tissue growth factor,46,47 a key molecule in the fibrotic process, which induces the ECM synthesis.17 It has been observed that increased expression of miR‑133 and miR‑30 may reduce connective tissue growth factor synthesis, leading to a decrease in collagen deposition.46
The profibrotic miRNAs include miR‑21 and miR‑125b (Table 1). MicroRNA‑21 is expressed exclusively in cardiac fibroblasts, and it can promote the proliferation and fibrosis of fibroblasts.30 Sygitowicz et al48 demonstrated an increase in miR‑21 expression in the serum of patients with symptomatic HF, independently of disease severity. The increase in miR‑21 expression did not correlate with the NT‑proBNP concentration but was significantly associated with an increase in the serum concentration of galectin 3, a biomarker of fibrotic process.48 Similar results were reported by Roy et al.49 They observed elevated miR‑21 expression in mouse hearts with myocardial ischemia–reperfusion. In a study performed by Jaffré et al,50 downregulated miR‑21 expression not only inhibited fibrosis but also prevented cardiac hypertrophy caused by thoracic aortic coarctation.
Pathological cardiomyocyte hypertrophy
Cardiomyocyte hypertrophy occurs in multiple pathological conditions, such as chronic overload, ischemic injury, and neurohormonal imbalance as part of an early adaptive reaction. However, this process may lead to cardiomyocyte death. Pathological concentric hypertrophy of cardiomyocytes is an independent risk factor and an indicator of poor prognosis in HF patients.37 Multiple molecular mechanisms are involved in the occurrence of pathological cardiomyocyte hypertrophy, especially the Ca2+-dependent pathway and the phosphatidylinositol 3‑kinase pathway.51,52 Multiple miRNAs can affect the Ca2+-dependent pathway. The miR‑1, miR‑9, miR‑26, and miR‑133 molecules exhibit protective effects against cardiac hypertrophy (Table 1).30 It was shown that miR‑1 inhibits cardiomyocyte hypertrophy in a rat model.30,51 In our previous study,48 we demonstrated a downregulated expression of circulating miR‑1 in patients with symptomatic HF. The degree of miR‑1 expression decreased with the severity of the New York Heart Association (NYHA) class. This effect was greater in patients in NYHA class IV than in those in NYHA class II or III. In our other studies,53,54 we also observed a significant negative correlation of reduced miR‑1 expression with the serum NT‑proBNP concentration and LV hypertrophy.
Myocardial cell apoptosis
Multiple miRNAs are associated with the regulation of myocardial cell apoptosis (Table 1). The key miRNA molecules involved in the induction of myocardial cell apoptosis are miR‑101a, miR‑200a, miR‑330 and miR‑320.30 It has been established that downregulated expression of miR‑200a leads to increased expression of β-catenin and cyclin D1 involved in the cell proliferation via activation of the E‑cadherin and Wnt/β-catenin signaling pathways.55 By inhibition of Akt phosphorylation, miR‑330 induces myocardial cell apoptosis,56 whereas miR‑101a may decrease the expression of Mcl‑1, an important protein of the antiapoptotic family of Bcl‑2.57 The proapoptotic action of miR‑320 is connected with its effects on the important cardioprotective heat‑shock protein 20.58 In contrast, the antiapoptotic effect is exerted by miRNA‑30, miRNA‑199, and miRNA‑206.30
Circulating microRNAs in the development of left ventricular remodeling and heart failure
Several studies have reported an association between circulating miRNA levels and the development of LV dysfunction and HF (Table 2).45,59-70 In this review, 21 dysregulated miRs were identified as potential biomarkers of LVR and HF development following AMI. The relationship between miRNA molecules frequently connected with these processes (including miR‑1, miR‑21, miR‑24, miR‑27a, miR‑29a/b, miR‑101, miR‑133a, and miR‑208a/b; Table 2) and cardiovascular diseases is well established. Some of these molecules are involved in pathological processes during HF development (Table 1). It is noteworthy that recent studies have demonstrated the role of other types of miRNAs (eg, miR‑34a and miR‑150) in postinfarction LVR. Among the selected types of miRNAs, there are also some novel biomarkers (eg, miR‑30a‑5p, miR‑1254) with limited data on their role in the pathogenesis of LVR and HF, although their prognostic value seems to be promising.
miRNAs | References |
Upregulated miRNAs | |
miR‑1 | Zile et al45
Grabmaier et al59 |
miR‑21 | Zile et al45
Liu et al60
Grabmaier et al59 |
miR‑29a | Zile et al45 |
miR‑29b | Grabmaier et al59 |
miR‑133a | Zile et al45 |
miR‑208a | Zile et al45 |
miR‑208b | Devaux et al61
Lv et al62
Lakhani et al63 |
miR‑499 | Devaux et al61 |
miR‑16 | Devaux et al64 |
miR‑27a | Devaux et al64 |
miR‑34a | Lv et al62
Lakhani et al63 |
miR‑146a | Liu et al60 |
miR‑155 | Latet et al65 |
miR‑184 | Liu et al66 |
miR‑126 | Lakhani et al63 |
miR‑1254 | de Gonzalo‑Calvo et al67 |
miR‑30a‑5p | Maciejak et al68 |
Downregulated miRNAs | |
miR‑150 | Devaux et al69
Devaux et al64
Lin et al70 |
miR‑101 | Devaux et al64 |
miR‑24 | Lakhani et al63 |
miR‑29a | Lakhani et al63 |
Some miR‑29 family members (miR‑29a and miR‑29b) have been reported as prognostic biomarkers in the development of LVR and HF.45,59,63 The miR‑29 family appears to affect the ECM remodeling of the heart by activating the Wnt signaling pathway.71 Target prediction analysis revealed that mRNAs for miR‑29 were involved in cardiac fibrosis processes, encoding collagen isoforms type I alpha 1 and 2, collagen type III alpha 1, matrix metalloproteinase 2, and others.47,72,73 Furthermore, miR‑29 family members are also involved in the regulation of cell proliferation, differentiation, and apoptosis.73 van Rooij et al72 demonstrated that in a mice model and human cardiac tissue samples, the expression of all 3 members of the miR‑29 family (miR‑29a, miR‑29b, and miR‑29c) was reduced in the border zone of the infarct. Subsequent analysis performed by van Rooij et al72 and Kriegel et al73 also confirmed that the downregulated expression of miR‑29 is involved in the process of cardiac fibrosis. Research conducted on isolated mice cardiac fibroblasts with transforming growth factor has shown the downregulated expression of miR‑29a, miR‑29b, and miR‑29c.72,73 Tao et al74 showed that decreased miR‑29a expression may play an important role in cardiac fibrotic tissue due to the regulation of vascular endothelial growth factor‑A and mitogen activated protein kinase signaling pathway. The circulating level of miR‑29a was significantly reduced in AMI patients with low LV ejection fraction, indicating its essential role in the occurrence of unfavorable LVR after AMI.63 On the other hand, it may be hypothesized that the upregulation of the miR‑29a expression early after AMI may be associated with limitation or inhibition of cardiac fibrosis or with promotion of ECM degradation, leading to more extensive LVR.45 Grabmaier et al59 showed that the elevated expression of miR‑29b early after AMI and during follow‑up was associated with the antifibrotic effects. Thus, the upregulation of miR‑29b could reflect a favorable outcome with regards to adverse LVR in post‑AMI patients. However, the above mechanisms have to be confirmed in further large‑scale clinical and animal studies.
The prognostic significance of miR‑150 in postinfarction LVR has also been confirmed.64,69,70 However, its function in the heart is still poorly understood. miR‑150 can be actively secreted by monocytes under various conditions.75 It can also be involved in various protective mechanisms against myocardial injury, cardiac hypertrophy, or myocardial fibrosis. The cardioprotective function of miR‑150 was confirmed in a mice model of AMI, where it was shown to regulate monocyte accumulation.75 Other studies76,77 showed that the upregulated expression of miR‑150 in the heart may lead to inhibition of cardiac hypertrophy and fibrosis via regulating of serum response factor and transcription factor c‑Myb. Moreover, the potentially cardioprotective effects exerted by miR‑150 may be related to the direct suppression of proapoptotic genes: zinc‑binding transcription factor induced by ischemia and proinflammatory ATP receptor in cardiomyocytes.78 On the other hand, the downregulation of circulating miR‑150 expression is associated with hypertrophy, LV ruptures, and unfavorable LVR after AMI.64,69 miR‑150 is involved in LVR by inhibiting the expression of its target genes: C‑reactive protein and adrenergic receptor beta 1.69 The association between miR‑150 and sepsis was also observed, which suggests its relation with inflammatory processes.79 Due to the fact that inflammation is engaged in the development of postinfarction LVR, further studies are needed to determine the proinflammatory effect of miR‑150.
The overexpression of miR‑30a‑5p is related to LV dysfunction after AMI.68 Circulating miR‑30a‑5p can be a novel prognostic biomarker. In silico functional analysis has shown that target genes for miR‑30a‑5p are probably involved in metabolic pathways associated with cardiovascular pathogenesis (eg, Wnt signaling pathway, calcium modulating pathway, fibroblast growth factor signaling).68 Some studies demonstrated the upregulation of miR‑30a expression in patients with AMI, LV hypertrophy, and HF.80,81 Pan et al81 published results in which they presented that miR‑30a is involved in cardiomyocytes autophagy by regulating its target gene, beclin 1. It was also shown that the decreased expression of miR‑30a‑5p and miR‑30c‑5p may be associated with the regulation of apoptosis in skeletal myoblast cells.82
Conclusions and future perspectives
MicroRNAs show a promising potential as unique molecular markers that more precisely illustrate the sequences of metabolic pathways leading from MI to HF than the current biomarkers used in clinical practice (such as natriuretic peptides or galectin 3). Hence, there is a need to identify the different types of miRNA molecules involved in the initiation and development of the disease. The current knowledge seems to suggest that miR‑29a, miR‑29b, miR‑150, and miR‑30a‑5p may be useful molecular markers in predicting the patient’s clinical status. The miRNA molecules presented in this review represent different groups of miRNAs, but all of them are involved in the regulation of basic processes such as cardiac fibrosis, apoptosis, autophagy, and inflammation associated with LV dysfunction after MI and HF. However, further clinical and animal studies are needed to confirm their pathophysiological function in the heart after MI as well as their usefulness in clinical practice, especially considering their potential role as an important therapeutic target in patients with developing HF.
- Dargie H. Heart failure post‑myocardial infarction: a review of the issues. Heart. 2005; 91 (suppl 2): ii3‑ii6. | Crossref
- Velagaleti RS, Pencina MJ, Murabito JM, et al. Long‑term trends in the incidence of heart failure after myocardial infarction. Circulation. 2008; 118: 2057‑2062. | Crossref
- Azevedo PS, Polegato BF, Minicucci MF, et al. Cardiac remodeling: concepts, clinical impact, pathophysiological mechanisms and pharmacologic treatment. Arq Bras Cardiol. 2016; 106: 62‑69. | Crossref
- Larina VN, Poryadin GV, Bogush NL, et al. Clinical profile of elderly patients with chronic heart failure in the presence of bendopnea. Pol Arch Int Med. 2019; 129: 939‑941. | Crossref
- Zornoff LA, Paiva SA, Duarte DR, Spadaro J. Ventricular remodeling after myocardial infarction: concepts and clinical implications. Arq Bras Cardiol. 2009; 92: 157‑164. | Crossref
ARTICLE INFORMATION