Extracellular vesicles in vascular pathophysiology: beyond their molecular content
Key words: biomarkers, ectosomes, exosomes, glycosylation, zeta potential



 CC BY-NC-SA 4.0
CC BY-NC-SA 4.0
Extracellular vesicles in vascular pathophysiology: beyond their molecular content
Extracellular vesicles (EVs) are released by all cells, both in physiological and pathological conditions. Their molecular charge and composition emerge as possible biomarkers, but EVs may also be considered for other clinical applications. This review discusses the role of other features of EVs, such as their lipid components or composition of glycans that form the EV corona and regulate EV biodistribution and uptake by target cells. The importance of EV electric charge has been discussed as a new insight into EV fate and destination.
Introduction
When about 50 years ago Wolf1 noticed that some tiny and highly abundant objects present in human blood plasma contribute to clotting events, it seemed that there is something beyond clotting factors that may support platelets / thrombocytes in their aggregation and formation of a thrombus. This coagulant particulate material released from platelets (referred to as “platelet dust”) was produced in considerably larger amounts than required for thrombin generation. It was detected not only in plasma but also in serum, and its presence seemed to be associated with the platelet‑like activity of the serum.1 The platelet activation leads to secretion of granules containing the procoagulant material and proteins involved in cytoskeletal arrangement, synaptic transportation, and secretion from their internal space.2 The processes of the formation and secretion of platelet “particles” are analogous to the processes occurring in other cells, including endothelial cells. They result in the formation of 2 types of membrane vesicles: bigger ones shed from the surface and named microvesicles (ectosomes) of 100 nm to 1 µm in diameter, and exosomes, measuring 40 nm to 100 nm in diameter. The latter are similar in size to the internal vesicles in multivesicular bodies (MVBs) and α-granules,3,4 and can be compared to endothelial vesiculation (Figure 1). Currently, the topic of extracellular vesicles (EVs) and their involvement in disease promotion and progression is gaining important insight in diagnostics and treatment.5 In this review, we focus on specific EV characteristics, still not intensively investigated, which contribute to the biological activity of EVs, that is, composition of the EV surface called a corona, and its charge.
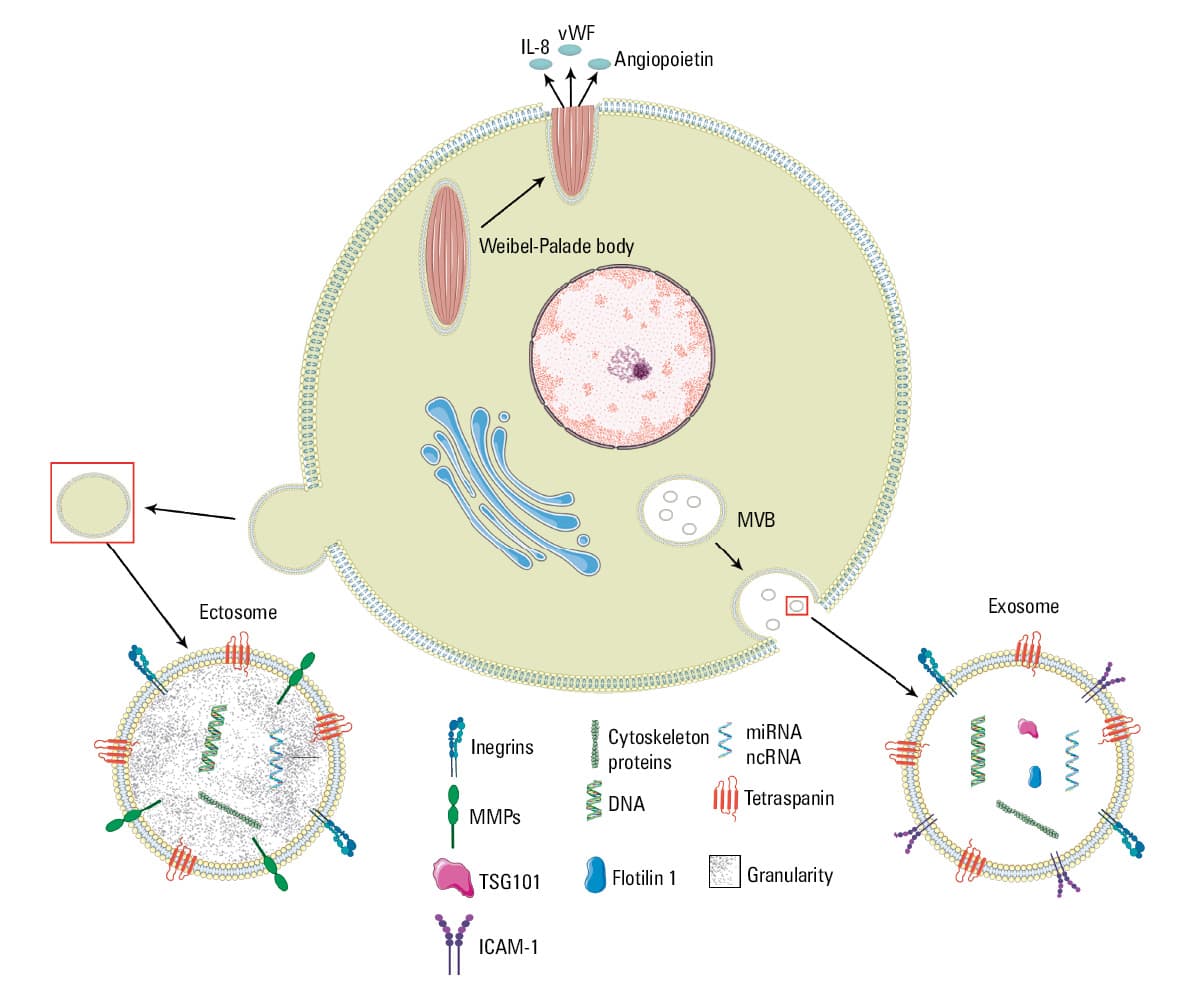
Abbreviations: IL‑8, interleukin 8; miRNA, micro‑RNA; ncRNA, noncoding RNA
The biological activity of EVs depends on their cargo and their biological availability and biostability. The estimated blood plasma concentration of EVs in healthy individuals is between 108 and 1014 EVs/ml. Such discrepancies, more than 6 orders of magnitude, depend mainly on the isolation protocol and possible contamination by other colloidal particles present in the plasma, such as lipoproteins and large protein aggregates.6 In total platelet poor plasma, the average platelet microvesicle (PMV) content has been estimated at 109 to 1010 EVs/ml in patients on antiplatelet drugs and healthy individuals. PMV abundance depends mainly on the plasma purity and preanalytical handling.7-11 Assuming that one‑third of the plasma EVs is of platelet origin, the number of the plasma EVs can be approximately 1010 EVs/ml.6 The average plasma residence time of intravenously delivered EVs ranges from 30 to 80 minutes and it is mainly regulated by the phagocyting activity of the mononuclear phagocyte system and by continuous turnover of EVs secreted by cells.12,13 Phagocytosis is one of the proposed mechanisms of EV internalization.14,15 Alternatively, EVs can be internalized by target cells in a variety of endocytic pathways (eg, clathrin‑dependent endocytosis16 and clathrin‑independent pathways, such as macropinocytosis,14 lipid raft‑mediated internalization, or caveole‑mediated uptake).14,17-19
What do extracellular vesicles contain?
The term EVs is used to refer to all membrane vesicles constituting a population of very diverse vesicular structures of different size and molecular content3,4,20 (Figure 2). Their molecular cargo has been intensively investigated and gathered in the biggest and manually curated compendia of molecular data for protein, lipid, and RNA, known as Vesiclepedia or ExoCarta21-23 or extracellular vesicle–associated DNA database (EV‑ADD).24 Both populations of EVs (ectosomes and exosomes) contain or carry specific proteins, which can be considered potential biomarkers.25-27 For the endothelial cells, the most pathognomonic proteins are urokinase plasminogen activator surface receptor (uPAR),28,29 von Willebrand factor (vWF),30 heat shock proteins,31 and metalloproteinases.32,33
Another molecular hallmark of EVs is microRNA (miRNA), specific, noncoding short RNA molecules.34,35 For years, EV miRNAs have been considered as a way of cell‑to‑cell communication. Recently, stoichiometric studies of miRNAs and exosomes have showed that most individual exosomes do not carry biologically significant amounts of miRNAs and they are unlikely to function as vehicles for miRNA‑based communication.36,37 Currently, EVs are rather considered a system for removing waste from the cells, which seems to be a good alternative to traditional laboratory biomarkers,38-40 or as vehicles for drug delivery systems for further clinical use.41,42
Molecular composition of EVs differs markedly also in terms of carbohydrates (glycans) attached to the surface proteins and forming a hydrophobic sugar overcoat (Figure 3). The presence and structure of glycans play a crucial role in cellular life and functioning of glycoproteins, for example, in cell‑cell recognition, pharmacokinetics, physical stability, and immunogenicity. Glycans attached to proteins exert various important biological functions, such as: 1) targeting recognition, 2) modulating protein activity, or 3) stabilizing protein folding. Changes in the glycoproteome probably contribute to the age‑related functional decline of the cardiovascular system as well as the heart and the aorta performance.43,44 Glycans covering the surface of EVs form a corona that has been recognized as a crucial mediator of EV functions.45,46 Glycomic profile of melanoma‑derived ectosomes showed important correlation with melanoma malignancy.47 Differences in the surface glycosylation pattern, particularly in N‑acetylglucosamine, mannose, and fucose‑binding lectins result in facilitated EV‑cell interactions and functional activation of endothelial cells.48 Glycans are key players in the regulation of EV uptake, through charge‑based effects or direct glycan recognition by targeting receptors.49
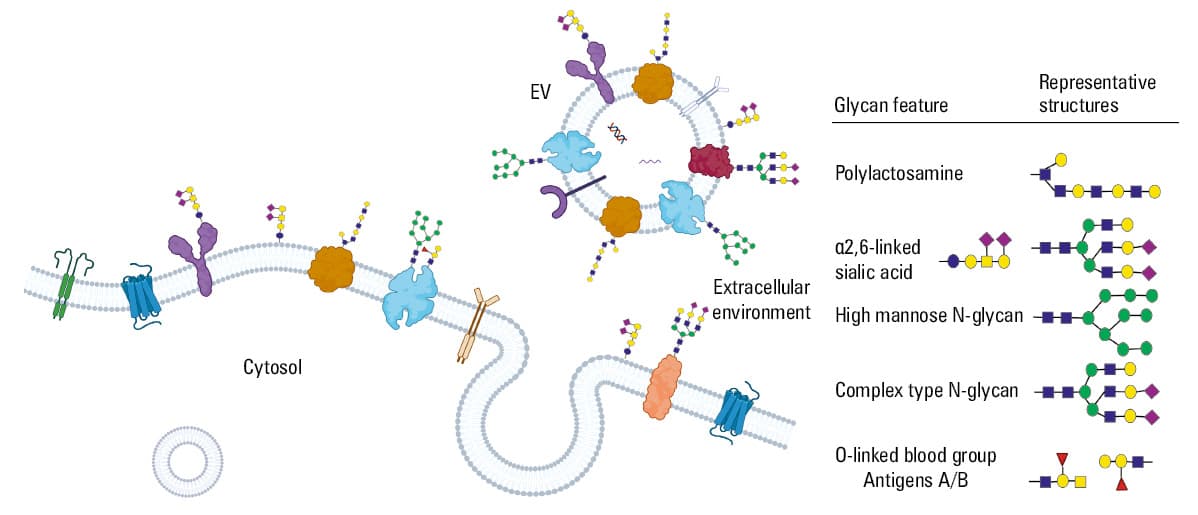
The importance of protein glycosylation for the biotechnology industry is highlighted by the fact that approximately 70% of therapeutic proteins, approved or in (pre-)clinical studies, are glycoproteins.
New feature of extracellular vesicles: a corona charge
Modification of surface N‑glycans increases EV uptake and reduces EV charge, expressed as the zeta potential (ZP), from negative toward neutral. The other contributors to the surface charge are phospholipids, the most common of which is a negatively charged phosphate group (PO43−). This group, when covalently bound to the lipid glycerol moiety of a 2‑chain fatty acid, forms the main group of charged fatty acids. Formation of EVs is very closely associated with the exposure of a membrane phospholipid phosphatidylserine (PS). Under normal conditions, PS is usually present in the inner membrane leaflet, but during EV secretion PS is transferred to the outer membrane leaflet.51 PS is composed of a negatively charged phosphate group attached to the serine at the hydroxyl end.
ZP is the measure that indicates the accumulation of negatively charged phospholipids in the inner membrane leaflet. Such accumulation of ions generates the ZP with an effective range of approximately 1 nm (Figure 4). Positively charged ions are attracted to the anionic surface, which is especially noticeable at the inner leaflet of the plasma membrane.
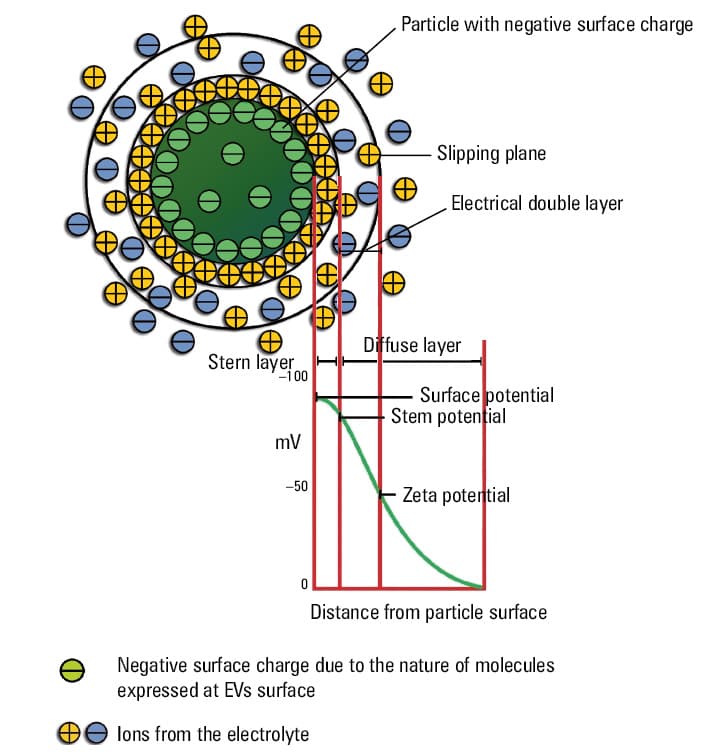
ZP is the electrostatic potential present at the boundary between the diffuse layer and the compact layer (also known as the raft layer) of a colloid system, in the case of EVs, it is an EV suspension in a body fluid. This potential is related to the surface charge of the EVs and is used as an indicator of their stability and ability to form aggregates. The more negative the ZP, the weaker the forces attaching EVs together, and the stronger the EV affinity to positive or less negative surfaces. The cell surface charge varies between positive and negative electric state, and it depends on the balance between negatively and positively charged molecules. In physiological conditions, the surface charge of endothelial cells is less negative,52 and it changes toward higher values in pathological conditions or during cancerogenesis.53-55
ZP as an indicator of colloidal stability of dispersed particles is influenced by the charge of a colloid surface. ZP is one of the useful measures to characterize colloidal stability, including EV aggregation and their electrophoretic mobility.56 Biological membranes (including EVs) bear negatively charged glycoproteins and glycolipids forming a complex corona with the surrounding medium and regulating biological properties, such as adhesiveness and internalization.57,58 The charge of the EV corona depends on different factors that control the interactions between the particle surface and the medium, such as pH and ionic strength of the medium56 (Figure 4).
As shown in Table 1, values of EV ZP differ surprisingly, ranging from –20 to –10 mV for EVs isolated from macrophages, astrocytes, and neurons57 or from –40 to –30 mV for EVs isolated from cancer cells or erythrocytes.54,58 These variations in ZP have not been explored yet, and may result from diverse separation methods, contamination of EVs with different colloids including lipoproteins, as well as external conditions, such as low pH, high ionic strength, or valency of surface cations.29,30
Origin | EV isolation methodology | EV zeta potential |
Abbreviations: AEx, adipose stem cells–derived exosomes; ASCs, adipose stem cells; DMSO, dimethyl sulfoxide; Exo‑5‑FU, 5‑fluorouracil‑loaded exosomes; Exo‑PAC; paclitaxel‑loaded exosomes; HLSC, adult human liver stem cells; iPS, induced pluripotent stem cells; MLC, mixed lymphocyte culture; MWCO, molecular weight cutoff; NB, neuroblastoma; PBS, phosphate‑buffered saline; RB, retinoblastoma; SEC, size exclusion chromatograhy; TEIR, Total Exosome Isolation Reagent for serum; TEx, tumor cell‑derived exosomes | ||
Adipose‑derived stem cells | 300 g, 10 min
2000 g, 10 min
10 000 g, 30 min
100 000 g, 70 min | −10.8 ± 0.65 mV59 |
Human serum | TEIR
ExoQuick
miRCURY
Ultracentrifugation (20 000 g, 30 min; 110 000 g, 70 min) | From –9.80 mV to –21.1 mV60 |
Bone‑marrow MSCs | 300 g, 5 min
16 500 g, 40 min
Filtration, 0.2 µm
120 000 g, 70 min | −30 ± 1.13 mV61 |
HLSC culture media
Human serum and saliva | 3000 g, 20 min
Filtration, 0.22 µm
3000 g, 20 min
10 000 g, 20 min
100 000 g, 60 min | HLSC culture medium –13.80 mV
Human serum –7.825 mV
Saliva –8.54 mV62 |
Glioblastoma cells | Exospin exosome purification kit | Empty exosome
−22.18 ± 8.73 mV
Incubation at 37 °C
−18.22 ± 1.23 mV
Sonication, −22.46 ± 0.63 mV63 |
Colon cancer HCT116 cell line and ASCs | 3000 g, 30 min
13 000 g, 70 min
Sample concentration using an Amicon Ultra‑15 Centrifugal Filter Devices (100 kDa, Millipore, Merck KGaA, Darmstadt, Germany)
120 000 g, 70 min
Filtration, 0.22 µm | TEx –9.20 ± 0.41 mV
AEx –7.22 ± 0.60 mV64 |
Lung cancer cells | 750 g, 15 min
2000 g, 20 min
Filtration, 0.45 µm
10 000 g, 45 min
Filtration, 0.22 µm
100 000 g, 90 min | −14.4 ±3.3 mV at RT for exosomes diluted in PBS with trehalose
−11.8 ±1.5 mV at RT for exosomes diluted in PBS with DMSO65 |
Human NB cell lines HTLA‑230, IMR‑32, SH‑SY5Y, and GI‑LI‑N | 300 g, 10 min
10 000 g, 30 min
Filtration, 0.22 µm
Filtration, 0.1 µm
100 000 g, 75 min
Washing in PBS
2 × ultracentrifugation 100 000 g, 75 min | HTLA‑230 −12.1 ±0.17 mV
IMR‑32 −14.8 ±1.55 mV
SH‑SY5Y −13.2 ±1.1 mV
GI‑LI‑N −12 ±0.15 mV66 |
Human umbilical cord–derived mesenchymal stromal cells | 3200 g, 30 min
Filtration, 0.2 µm
Sample concentration using an Amicon Ultra‑15 filter unit with Ultracel‑100 membrane (MWCO = 100 kDa, Merck Millipore, Cat. No. UFC910024)
SEC column | From −7.73 ±3.76 mV to −12.4 ±2.5 mV67 |
Human urine–derived stem cells | Centrifugation using an Amicon Ultra15 Centrifugal Filter Tube (10 kDa; Millipore)
ExoQuick‑TC Solution (System Biosciences, Palo Alto, California, United States)
1500 g, 30 min | −2.02 ±0.03 mV68 |
Red blood cells | 1500 g, 10 min
3000 g, 15 min
25 000 g, 60 min
~200 g, 120 min | −36.4 ±7.8 mV69 |
Raw bovine milk | 13 000 g, 30 min
90 000 g, 60 min
180 000 g, 120 min
Filtration, 0.2 µm | Exo‑PAC −28.28 ±1.8 mV
Exo‑5‑FU −27 ±1.6 mV; plain exosomes −23 ±1.2 mV70 |
Human serum | Total exosome isolation from serum (Invitrogen by Thermo Fisher Scientific, Vilnius, Lithuania) | Serum small EVs (sEVs) from RB individuals −11.04 ±0.4 mV
Serum small EVs (sEVs) from non‑RB individuals –12.72 ±1.7 mV71 |
Murine cardiac fibroblasts (CF) and CF‑derived iPS | 3000 g, 10 min
Filtration, 0.2 µm
precipitation overnight in PEG buffer at 4 ˚C
1500 g, 30 min | CF exosomes −14.22mV
iPS exosomes −15.44 mV72 |
Human immortalized microvascular endothelial cell line (TIME) | 2000 g, 30 min
18 000 g, 30 min
150 000 g, 90 min | Ectosomes –9.3 ±0.7 mV
Exosomes –11.35 ±1.9 mV22 |
Concluding remarks
EVs are very attractive research objects as potential biomarkers of various pathologies due to their specific cargo and abundance in the cellular environment. As they have different cellular origin, they also differ in their metabolic competencies, and carry a variety of bioactive molecules, such as miRNAs, proteins, and lipids. However, their biodistribution and availability are also important and still underestimated features. For their further use in clinical practice as potential drug carriers, their biodistribution and uptake by target cells should be considered. To better understand the mechanisms of cell internalization, most attention should be given to the EV surface properties related to the EV corona and surface charge, characterized by glycosylation and ZP.
- Wolf P. The nature and significance of platelet products in human plasma. Br J Haematol. 1967; 13: 269‑288. | Crossref
- Kasprzyk J, Stępień E, Piekoszewski W. Application of nano‑LC‑MALDI‑TOF/TOF‑MS for proteomic analysis of microvesicles. Clin Biochem. 2017; 50: 241‑243. | Crossref
- Heijnen HF, Schiel AE, Fijnheer R, et al. Activated platelets release two types of membrane vesicles: microvesicles by surface shedding and exosomes derived from exocytosis of multivesicular bodies and alpha‑granules. Blood. 1999; 94: 3791‑3799. | Crossref
- Cocucci E, Meldolesi J. Ectosomes and exosomes: shedding the confusion between extracellular vesicles. Trends Cell Biol. 2015; 25: 364‑372. | Crossref
- Stępień E, Rząca C, Moskal P. Novel biomarker and drug delivery systems for theranostics ‑ extracellular vesicles. Bio‑Algorithms and Med‑Syst. 2021; 17: 301‑309. | Crossref
ARTICLE INFORMATION