Epstein–Barr virus and immune chaos: the link between reactivation and Toll-like receptor dysregulation in immunodeficiency
Key words: chronic lymphocytic leukemia, common variable immunodeficiency, Epstein–Barr virus, reactivation, Toll-like receptor

 CC BY 4.0
CC BY 4.0
Epstein–Barr virus and immune chaos: the link between reactivation and Toll-like receptor dysregulation in immunodeficiency
Introduction: The immune system protects the body against pathogens, and its dysfunction leads to primary and secondary immunodeficiencies, increasing infection susceptibility. Epstein–Barr virus (EBV) reactivation is linked to immune homeostasis disorders, particularly in common variable immunodeficiency (CVID) and chronic lymphocytic leukemia (CLL). Toll‑like receptor (TLR) pathways play a crucial role in innate immunity, and their deregulation may contribute to immune dysfunction.
Objectives: This study aimed to assess the impact of EBV reactivation on immune homeostasis, focusing on TLR2, TLR4, TLR7, and TLR9 expression in T and B lymphocyte subpopulations and their soluble forms in CVID and CLL patients.
Patients and methods: The study included 60 CVID patients, 60 CLL patients, and 30 healthy controls. EBV antigens, viral DNA levels, T and B lymphocyte immunophenotypes, and serum soluble TLR (sTLR) concentrations were analyzed using flow cytometry and enzyme‑linked immunosorbent assay.
Results: EBV reactivation was detected in 55% of CVID and 60% of CLL patients. These patients showed significant TLR expression disturbances and increased sTLR levels. Notably, TLR7 and TLR9 expression was elevated in CD4+, CD8+ T, and CD19+ B cells, correlating with EBV load and immune dysfunction severity.
Conclusions: EBV reactivation plays a key role in TLR pathway deregulation in CVID and CLL, potentially accelerating disease progression and increasing infection risk. TLR expression and sTLR levels could serve as biomarkers for EBV reactivation, aiding therapeutic strategies to stabilize immune responses.
What's new?
This study provides a comprehensive and detailed analysis of the impact of Epstein–Barr virus (EBV) reactivation on Toll‑like receptor (TLR) signaling in the context of both primary and secondary immunodeficiencies, particularly in common variable immunodeficiency and chronic lymphocytic leukemia patients. In addition to confirming the association between EBV reactivation and immune dysregulation, our findings highlight that disruptions in TLR signaling may be a mechanism contributing to the immune imbalance in these immunocompromised individuals. A key advance in this research is the characterization of the percentages of TLR2, TLR4, TLR7, and TLR9 in different T- and B‑cell subpopulations, showing significant changes associated with EBV reactivation. Furthermore, this study is the first to comprehensively assess soluble forms of TLR in serum in this context, providing new insights into their potential role as biomarkers of EBV‑induced immune dysfunction.
Introduction
The immune system is a key defense mechanism against a variety of pathogens, playing a vital role in maintaining overall health and well‑being. Dysregulation of immune responses can result in immune deficiencies, making individuals susceptible to recurrent and severe infections. Epstein–Barr virus (EBV), a member of the Herpesviridae family, is a ubiquitous virus that infects more than 90% of the world’s population. Primary EBV infection is often asymptomatic or causes infectious mononucleosis.1 After primary infection, the virus establishes latency in B lymphocytes and can reactivate periodically throughout the life of the individual.2-5 EBV reactivation is associated with various complications, including lymphoproliferative disorders and autoimmune or neoplastic diseases.6,7 It is also involved in exacerbating immune dysregulation in patients with primary (PID) and secondary immunodeficiencies (SID).8-13 However, the mechanisms of its interaction with the immune system in the course of PIDs and SIDs are not yet fully understood. Dysregulation of Toll‑like receptor (TLR) signaling pathways, key components of the innate immune system, has emerged as a potential factor contributing to immune imbalance in these individuals.14 TLRs are a class of pattern recognition receptors that recognize conserved molecular patterns present in many pathogens, including viruses. Activation of TLR triggers a signaling cascade that initiates immune responses, leading to the production of proinflammatory cytokines, chemokines, and the activation of antigen‑presenting cells. Dysregulation of TLR signaling pathways can disrupt immune homeostasis, contributing to immune deficiencies and autoimmune diseases.15-18 Understanding the impact of EBV reactivation on immune stability, with particular emphasis on the dysregulation of TLR signaling pathways, is of great importance to elucidate the mechanisms underlying immune dysfunction in patients with immunodeficiencies. Therefore, this study aimed to investigate the impact of EBV reactivation on immune stability in patients with selected PIDs and SIDs, with particular emphasis on the dysregulation of TLR signaling pathways. By assessing viral reactivation and TLR expression levels on selected T- and B- cell subpopulations, as well as studying their interactions, we can enhance our understanding of the immunopathogenesis of immunodeficiencies associated with EBV reactivation and TLR dysregulation, paving the way for the development of targeted therapeutic interventions to restore immune stability and improve clinical outcomes in affected individuals.
Patients and methods
Research material and characteristics of patients
The study included 60 individuals with newly diagnosed common variable immunodeficiency (CVID) and 60 individuals with chronic lymphocytic leukemia (CLL), selected based on specific criteria established by a clinical immunology specialist and / or a hematologist.19,20 In the CLL diagnostics, lymphocytosis (≥5 × 10⁹/l) persisting for 3 months, confirmed by flow cytometry (CD5, CD19, CD23, low CD20, and immunoglobulins) was found. Other lymphoproliferative diseases were excluded. Additionally, hypogammaglobulinemia, recurrent infections, and clinical symptoms, such as enlarged nodes and spleen, were noted. In CVID, a decrease in immunoglobulin (Ig) G, IgA, and / or IgM and a lack of response to vaccination were confirmed. SIDs were excluded. Recurrent infections, splenomegaly, and enteropathies were identified. Participants had to be 18 years or older, have a life expectancy of at least 12 months, and abstain from immunosuppressive treatments for at least 3 months before the study. Informed consent was required. Exclusion criteria comprised active viral, bacterial, or fungal infections (SARS‑CoV‑2, influenza A and B viruses, respiratory syncytial virus, adenoviruses, rhinoviruses, hepatitis B virus, hepatitis C virus, hepatitis A virus, HIV, cytomegalovirus, EBV, herpes simplex virus type 1 and 2, and rotaviruses and noroviruses. Bacterial infections, such as Streptococcus pneumoniae, Haemophilus influenzae, Mycoplasma pneumoniae, Legionella pneumophila, Escherichia coli, Klebsiella spp., Proteus spp., Staphylococcus aureus, Enterococcus spp., Salmonella spp., Shigella spp., Clostridium difficile or Treponema pallidum were not detected. Fungal diagnostics excluded the presence of Candida spp., Aspergillus spp., Cryptococcus neoformans, Pneumocystis jirovecii, and Histoplasma capsulatum. The remainig exclusion criteria comprised severe allergies, recent hematopoietic cell or organ transplants, active malignancies, autoimmune diseases, pregnancy, lactation, and participation in clinical trials. Individuals with central nervous system metastases or mental illness were also excluded. A control group of 30 healthy volunteers who met the same inclusion and exclusion criteria as the patient groups was recruited. Median (interquartile range [IQR]) age of the CVID group was 46.1 (27–59) years, and the group included 28 women and 32 men. Median (IQR) age of the CLL group was 65.2 (51–73) years, and the group comprised of 12 women and 48 men. Median (IQR) age of the healthy volunteers was 56.8 (32–75) years, and the group consisted of 17 women and 13 men.
A 10‑ml blood sample was collected to analyze hematological parameters, biochemical markers, and the immunophenotyping of T and B lymphocytes expressing TLRs, while an additional 5 ml was used for serum analysis to measure soluble TLR concentrations and anti‑EBV antibodies.
Immunophenotyping
Immunophenotyping of peripheral blood lymphocytes was performed using flow cytometry. Blood samples were labeled with a panel of monoclonal antibodies (including anti‑CD4, CD3, CD8, CD19, CD45, and antibodies against TLR2, TLR4, TLR7, TLR8, TLR3, and TLR9). After staining, erythrocytes were removed with a lysis buffer, and cells were washed and analyzed on a CytoFLEX LX instrument (Beckman Coulter, Brea, California, United States). Data were analyzed using Kaluza software (Beckman Coulter), and quality was controlled daily using CytoFLEX QC Fluorospheres (Beckman Coulter). An example gating strategy is shown in Supplementary material, Figure S1.
Quantification of soluble Toll‑like receptor forms and serological profiles of anti‑Epstein–Barr virus antibodies
To assess the concentration of soluble TLR forms, enzyme‑linked immunosorbent assays (ELISA) were performed on serum samples from all study participants. Commercially available kits were used, including the Human TLR2, TLR3, TLR4 ELISA Kits (Abcam, Cambridge, United Kingdom), and TLR7, TLR8, TLR9 ELISA Kits (MyBiosource, San Diego, California, United States). Measurements were taken using the VictorTM3 reader (PerkinElmer, Waltham, Massachusetts, United States).
In addition, a qualitative assessment of specific anti‑EBV antibodies (IgA, IgM, IgG) was conducted in both the study and control groups. The antibodies were detected using ELISA kits targeting viral‑capsid antigen (VCA), early antigen (EA), and Epstein–Barr nuclear antigen 1 (EBNA‑1). Serological criteria for EBV infection reactivation are based on the presence of EA IgG antibodies which are the main marker of active virus replication. Additionally, the presence of VCA IgG (++), VCA IgA (+/–) and variable expression of VCA IgM (+/–) and EA IgA (+/–) are observed. The EBNA‑1 IgG (+/–) result is also of diagnostic value, the absence or variable level of which differentiates EBV reactivation from previous infections.21 The assays were performed according to the manufacturer’s instructions (Demeditec Diagnostics GmbH, Kiel, Germany), and absorbance was measured using the VictorTM3 microplate reader (Perkin Elmer, Waltham, Massachusetts, United States). Antibody levels were expressed in U/ml, with values above 11 considered positive.
Evaluation of the number of Epstein–Barr virus copies in the genetic material from peripheral blood mononuclear cell
The EBV DNA copy number was determined using the ISEX EBV polymerase chain reaction (PCR) kit (GeneProof, Brno, Czech Republic). Each sample was tested twice with a negative control, and the EBNA‑1 gene sequence was amplified using the 7300 Real‑Time PCR System (Thermo Fisher Scientific, Waltham, Massachusetts, United States). The results were adjusted for DNA isolation efficiency and expressed as viral DNA copy number per µg of DNA. Samples with fewer than 10 copies/µl were classified as negative for EBV.
Statistical analysis
The data obtained from the study were analyzed using Tibco Statistica 13.3 software (Tibco Software Inc., Palo Alto, California, United States). The normality of the data distribution was determined using the Shapiro–Wilk test. Differences between the groups were analyzed using the Kruskal–Wallis test, followed by the Dunn post hoc test. The Bonferroni method adjusted the P values for the Dunn test for multiple comparisons. Spearman correlation coefficients were utilized to study the relationships between variable pairs. Receiver operating characteristic (ROC) curves were used to determine the diagnostic performance of the laboratory test for patient‑related parameters. To visualize the obtained data clearly and concisely, GraphPad Prism v. 9.4.1 was used (GraphPad Prism Software, San Diego, California, United States). For all statistical tests, a P value below 0.05 was considered significant.
Ethics
The study protocol received the necessary approval from the Bioethics Committee of the Medical University of Lublin (KE‑0254/186/06/2023).
Results
Our analysis included 150 participants: 60 with CVID, 60 with CLL, and 30 healthy controls. The CVID and CLL patients were newly diagnosed and had not received any immune‑affecting treatments. Median time from symptom onset to diagnosis was 5.5 years for CVID and 37 months for CLL (IQR, 25–49). The CLL patients were in Rai stage 0 (n = 37) and I (n = 23). In the 12 months preceding diagnosis, both the CVID and CLL patients had multiple episodes of infections requiring antibiotic therapy, mainly involving the upper and lower respiratory, gastrointestinal, and urinary tracts. In the CVID group, 20% of the patients had 3–5 infection episodes, 36.67% had 6–8 episodes, 26.67% had 9–11 episodes, and 16.66% had more than 12 episodes. Among the CLL patients, 8.33% had 3–5 episodes, 28.34% had 6–8 episodes, 33.33% had 9–11 episodes, and 30% had more than 12. Among the healthy volunteers, 30% reported 1–2 infections, while the remaining 70% did not exhibit infection symptoms in the past year. In the initial stage of the study, hematological parameters, immunophenotype, TLR expression on T and B lymphocytes, and soluble TLR (sTLR) levels were analyzed. These data (Supplementary material, Table S1) showed significant differences between the immunodeficient patients and the controls, as well as between the CVID and CLL groups, leading to further investigation of EBV reactivation’s role in immune dysregulation.
The next stage of the study was the analysis of the level of antibodies against EA, VCA, and EBNA‑1 antigens in the patients diagnosed with CVID and CLL and the healthy volunteers. It was performed to assess EBV reactivation. The results are presented in Supplementary material, Table S2. EBV reactivation was observed in 55% of the CVID patients and 60% of the CLL patients, labeled as EBV+. No reactivation was detected in the remaining patients and the healthy volunteers, marked as EBV–, although previous EBV exposure was confirmed. Additionally, EBV viral load in peripheral blood mononuclear cells was measured in reactivated patients, showing a 30% higher mean viral load in the CLL patients, compared with the CVID patients. No viral load was detected in the remaining participants (below the test detection limit of 10 copies).
Effect of Epstien–Barr virus reactivation on hematological, biochemical, and immunophenotypic parameters, as well as infection frequency in the patients with common variable immunodeficiency, chronic lymphocytic leukemia, and the healthy volunteers
Due to the high rate of EBV reactivation and increased viral load in the CVID and CLL patients, we analyzed peripheral blood parameters (Supplementary material, Table S3) and immune cell populations (Supplementary material , Table S4) in terms of EBV reactivation. The analysis of hematological parameters showed significant differences among all 3 groups, especially in the context of EBV reactivation. The CVID EBV+ patients had lower leukocyte counts (6.02 × 10³/mm³ vs 6.43 × 10³/mm³; P <0.001), neutrophils, and monocytes, compared with the CVID EBV– population. They also showed lower erythrocyte, hemoglobin, and platelet counts, compared with the other groups. In the CLL group, increased lymphocyte counts (26.78 × 10³/mm³ in EBV+ vs 1.36 × 10³/mm³ in CVID EBV–; P <0.001) and leukocytosis predominated, regardless of the EBV status. Compared with the healthy volunteers (platelet count, 273.22 × 10³/mm³), the CVID and CLL patients showed lower platelet counts, especially in the CVID EBV+ groupd (109.85 × 10³/mm³; P <0.001). Anemia was more severe in the CVID EBV+ (hemoglobin, 8.96 g/dl) than in the CVID EBV– and CLL groups, and in both diseases, hemoglobin values were significantly lower than in the control group (13.97 g/dl) (Supplementary material, Table S3). In the analysis of the occurrence of immune system cell populations, we also noted several significant changes among the analyzed groups of patients (Supplementary material, Table S4). The increase in the percentage of CD3+, CD8+, and CD19+ lymphocytes in the patients with CLL EBV+, as compared with the CLL EBV– participants, deserves special attention. No significant changes were observed between the patients with CVID EBV+ and CVID EBV–. Compared with the healthy volunteers, the analysis of all patient groups with immunodeficiencies demonstrated significant differences in nearly all the examined hematological, biochemical, and immunophenotypic parameters of peripheral blood (Supplementary material, Tables S5 and S6). The frequency of infections requiring antibiotic therapy in the year preceding the diagnosis was higher in the patients with CVID EBV+, compared with those with EBV–. In the first group, 42.42% had 6–8 infections, 30.30% had 9–11 infections, while 27.28% had more than 12 infections during the year. Among the EBV– patients, the highest percentage (44.44%) had 3–5 infections, 29.63% suffered from 6–8 infections, 22.22% had 9–11 infections, and 3.71%, more than 12 infections during the year. The same analysis was performed for the CLL patients, who also exhibited an increased infection frequency. In the patients with EBV+, the data were as follows: 2.78% in the range of 3–5 infections, 25% in the range of 6–8 infections, 36.11% in the range of 9–11 infections, and 31.11%, more than 12 infections during the year. In the EBV– group, 16.66% had 3–5 infections, 33.33% had 6–8 infections, 29.17 suffered from 9–11 infections, and 20.83% contracted more than 12 infections during the past year.
The influence of Epstein–Barr virus reactivation on the percentage of Toll‑like receptors on selected immune cells
The analysis of selected populations of immune cells (especially T and B lymphocytes) in the patients with CVID and CLL showed significant changes in the percentage of T and B lymphocytes positively expressing selected TLRs (extracellular: TLR2, TLR4 and intracellular: TLR3, TLR7, TLR8, and TLR9) in all patient groups. Disturbances of the immune response via TLRs may contribute to the regulation of immune homeostasis of the whole organism and affect the proper antibacterial and antiviral response. This may indirectly contribute to the increased number of infections observed in patients with both PIDs and SIDs. By examining TLR expression patterns in EBV reactivation, we aimed to understand the potential impact of altered TLR signaling on the immune response against this particular viral infection. We also intended to comprehend its impact on immune homeostasis and the ability to generate effective immune responses against infections, especially in immunocompromised patients. Therefore, in the next step of our research, we decided to analyze the percentage of selected T- and B‑cell subpopulations positively expressing TLR2, TLR3, TLR4, TLR7, TLR8, and TLR9 in the patients with CVID and CLL in the context of EBV reactivation. The obtained test results are presented in Supplementary material, Table S7, and for better visualization of the occurring changes, also in Figures 1, 2, 3.
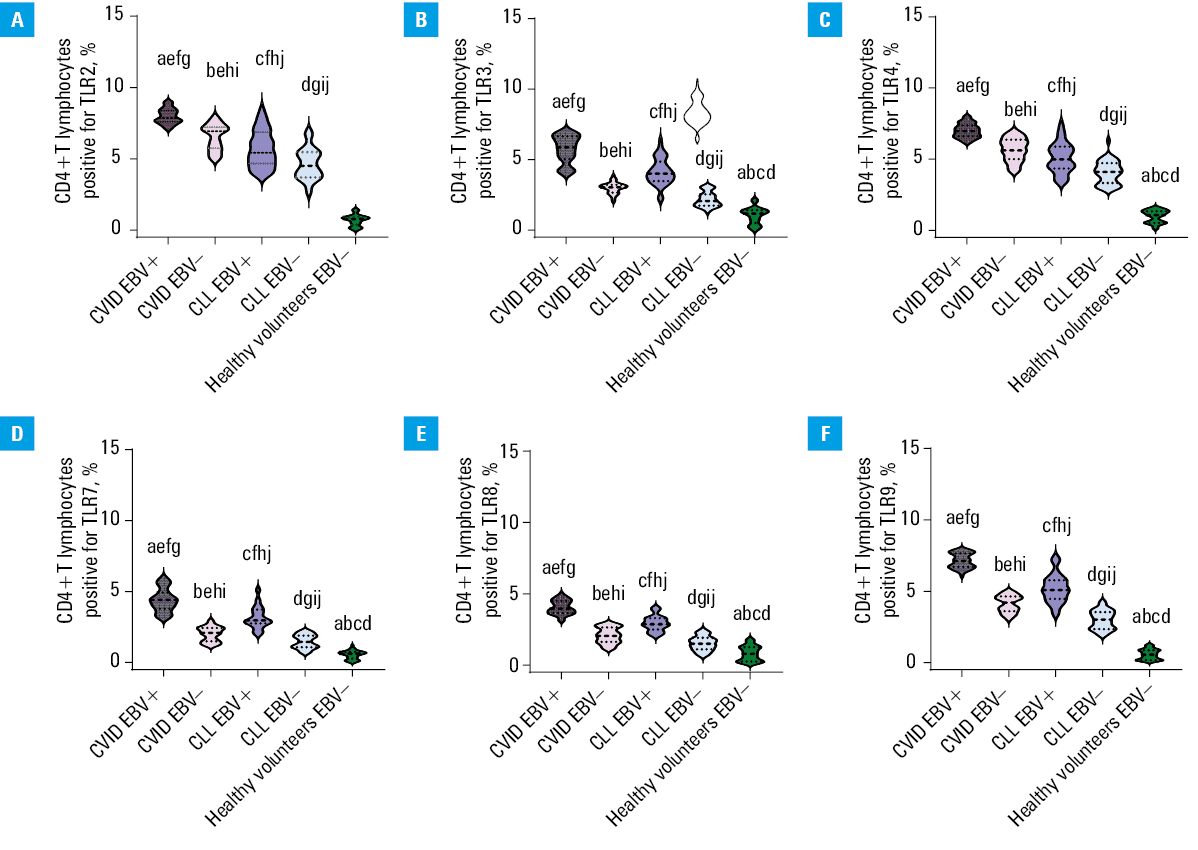
Abbreviations: CLL, chronic lymphocytic leukemia; CVID, common variable immunodeficiency; EBV, Epstein–Barr virus; TRL, Toll‑like receptor
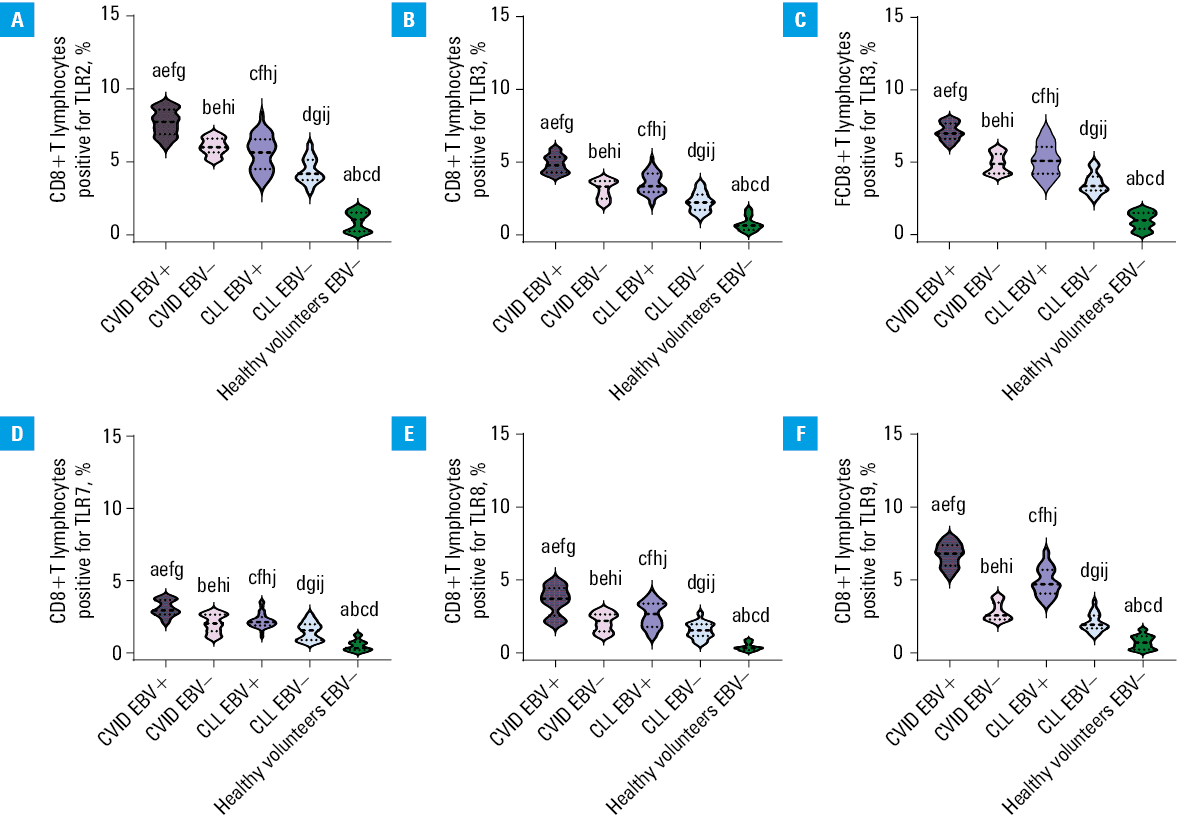
Abbreviations: see Figure 1
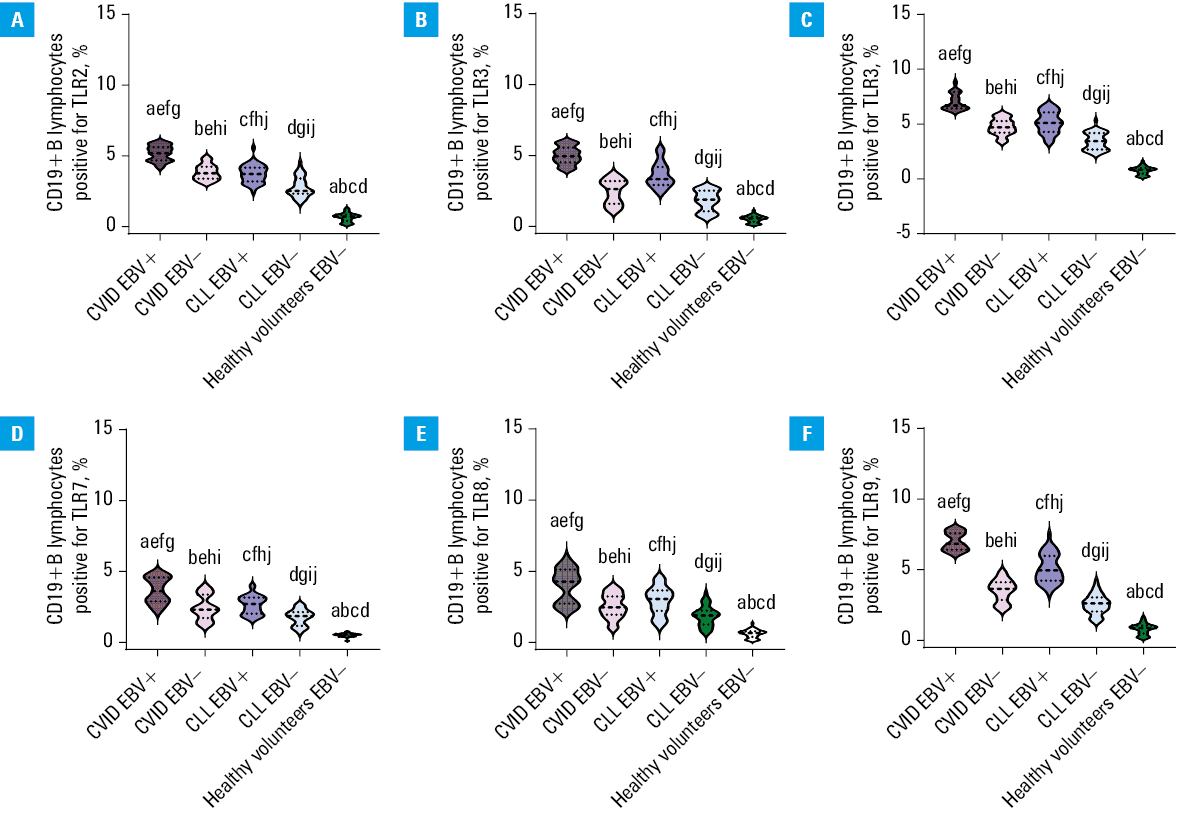
Abbreviations: see Figure 1
Based on the results, we showed that EBV virus reactivation significantly affected the deregulated activation of the immune response via TLRs. In both the CVID and CLL groups, the percentage of all analyzed T and B lymphocyte subpopulations, positive for all tested TLRs, was higher in the EBV+ than in the EBV– patients (Figures 1, 2, 3). Detailed analysis of the mean values of individual TLRs on T and B lymphocytes in the CVID EBV+ patients showed a significant increase, in comparison with EBV–. The largest observed differences were: CD8+TLR9+ (2.37‑fold), CD4+TLR7+ (2.19‑fold), CD19+TLR3+ (2.05‑fold), CD19+TLR9+ (1.98‑fold), CD4+TLR8+ (1.94‑fold), and CD4+TLR3+ (1.92‑fold). For the remaining analyzed expressions, the differences ranged from 1.22‑fold for CD4+TLR2+ to 1.73‑fold for CD4+TLR9+. Similar relationships were also observed between the patients with CLL EBV+ and CLL EBV–. Here, the largest differences were observed for: CD8+TLR9+ (2.34‑fold), CD4+TLR7+ (2.18‑fold), CD19+TLR3+ (2.03‑fold), CD19+TLR9+ (1.99‑fold), CD4+TLR8+ (1.93‑fold), and CD4+TLR3+ (1.9‑fold). For the remaining analyzed expressions, the differences ranged from 1.23 to 1.73‑fold. In addition, as compared with the CLL group, a significant increase in the expression of the examined TLRs was observed in the patients diagnosed with CVID, also in the case of detected EBV reactivation. As can be seen from the observed changes, the greatest differences in the percentage of occurrence of the studied lymphocyte subpopulations showed increased expression of mainly intracellular TLRs, responsible for the recognition of double‑stranded RNA by TLR3, single‑stranded RNA by TLR7 and TLR8, and CpG by TLR9, respectively. Statistical analyses concerning the comparison of the selected groups of patients with the healthy volunteers are presented in Supplementary material, Table S8.
The impact of Epstein–Barr virus reactivation on the serum concentration of soluble Toll‑like receptors in patients with immunodeficiencies
The next step in our analysis was to assess the effect of EBV reactivation on the serum concentration of sTLRs in the patients diagnosed with CVID and CLL, as well as the healthy volunteers. The obtained results are presented in Figure 4, and the details are presented in Supplementary material, Table S9.
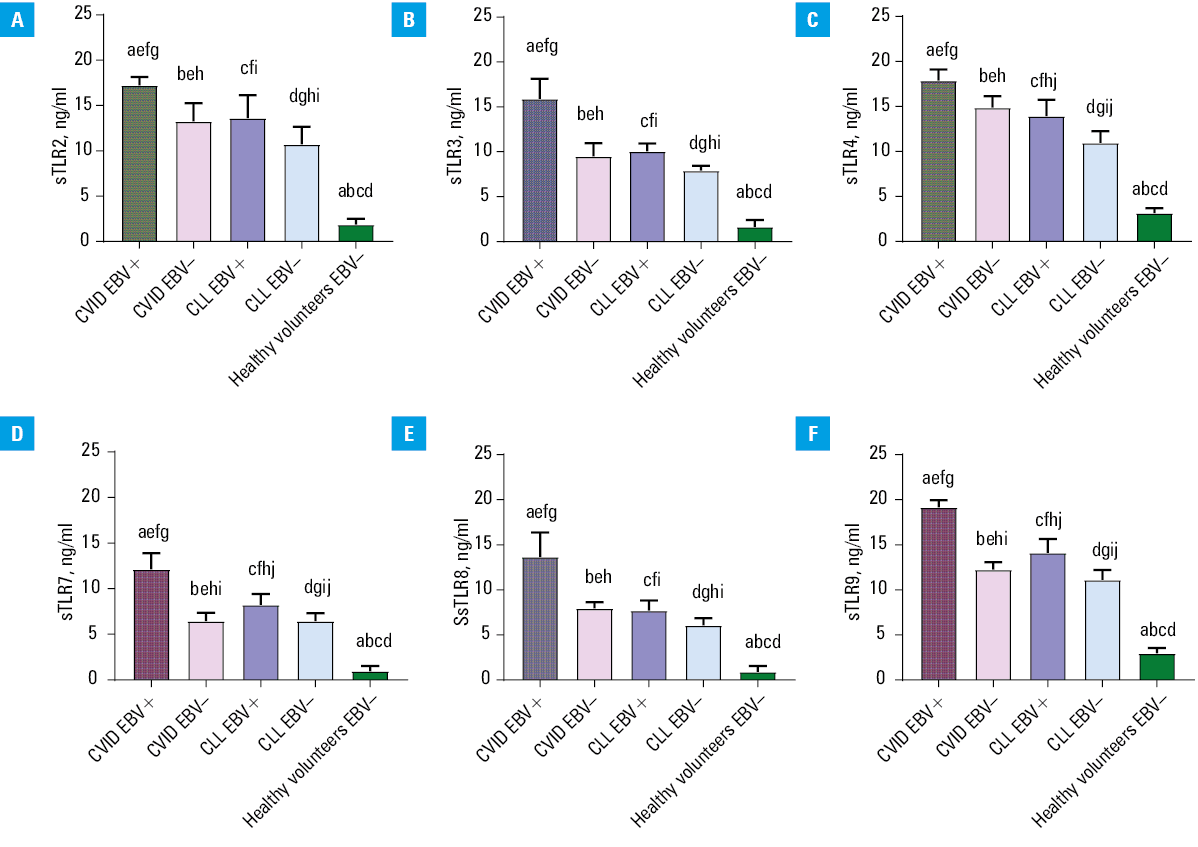
Abbreviations: sTLR, soluble form of Toll‑like receptor; others, see Figure 1
The relationships observed in the previous analyses were also evident in this analysis. This means that the patients with the EBV reactivation had higher concentrations of soluble forms of all the tested TLRs in the serum, compared with EBV– patients (both the CVID and CLL). Detailed analysis of mean serum sTLR values in the CVID EBV+ patients showed an increase in observed concentrations, compared with the CVID EBV– patients by: 1.2‑fold for sTLR4, 1.3‑fold for sTLR2, 1.56‑fold for sTLR9, 1.67‑fold for sTLR3, 1.71‑fold for sTLR8, and 1.88‑fold for sTLR7. This means that despite the lowest observed serum sTLR7 concentrations, its difference in the occurrence of EBV reactivation was the greatest. In the CLL patients, EBV reactivation affected serum concentrations of the studied sTLRs less variably, ranging from 1.26‑fold for sTLR2 to 1.33‑fold for sTLR7. Moreover, we showed that the CVID EBV+ patients had increased levels of all analyzed sTLRs, compared with the CLL EBV+ patients. Statistical analyses comparing the results of all patient groups are presented in Supplementary material, Table S10.
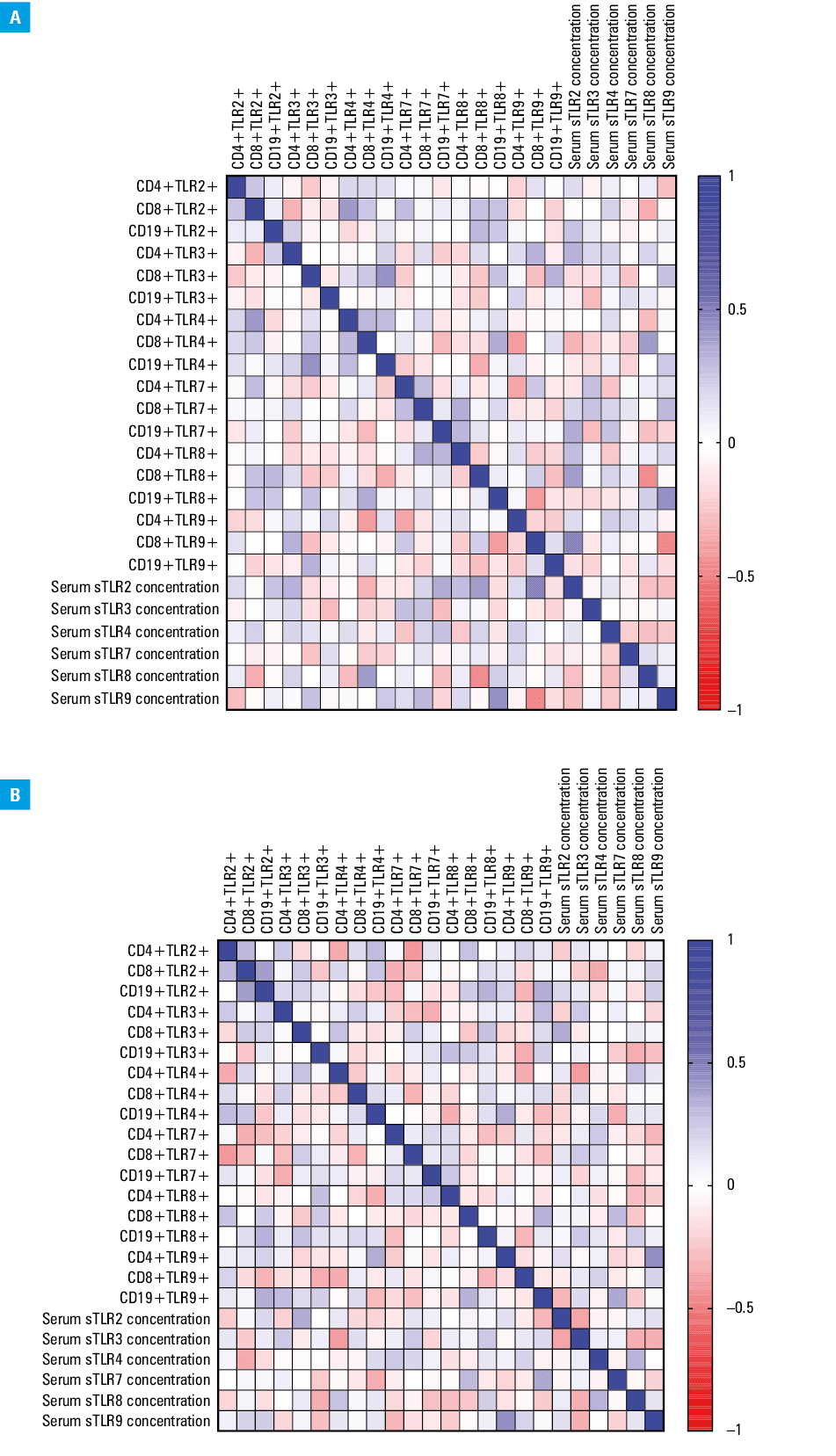
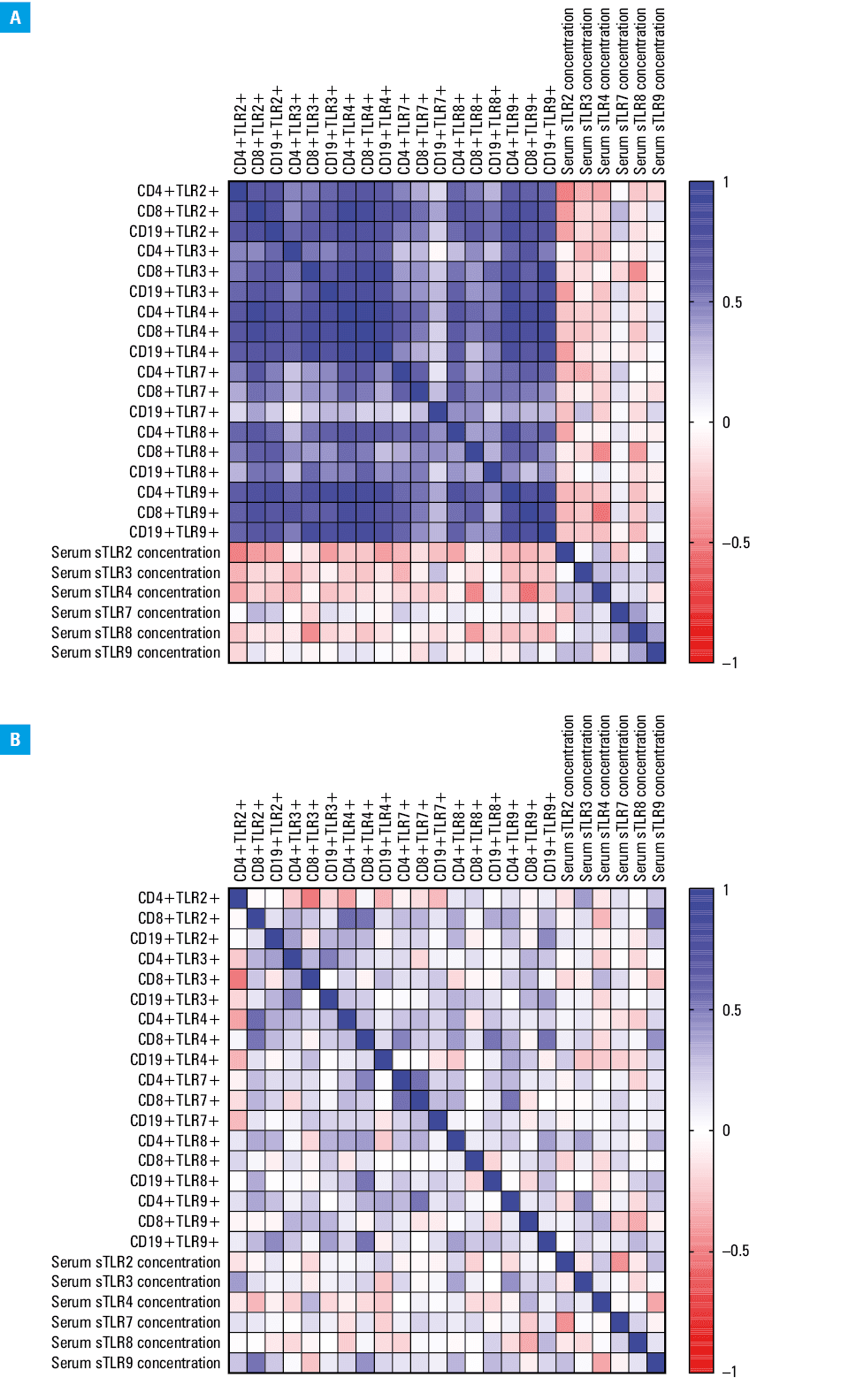
Correlation analysis of Epstein–Barr virus reactivation and immune system parameters in immunodeficient patients
Given the significant changes observed in the expression of TLRs and their soluble forms, a correlation analysis was conducted to explore the impact of EBV reactivation on immune system parameters in both the EBV+ and EBV– patients. For the CVID EBV+ patients, 20 significant correlations were identified, with 11 being negative (5 low, 6 moderate) and 8 positive (3 low, 4 moderate; Supplementary material, Table S11; Figure 5A). In the CVID EBV– patients, 18 significant correlations were found, including 10 negative (5 low, 5 moderate) and 8 positive (4 low, 4 moderate) (Supplementary material, Table S12, Figure 5B). In the CLL EBV+ patients, 156 significant correlations were observed, with the majority (n = 129) related to the changes in T and B lymphocyte populations expressing TLRs, all of which were positive (3 low, 53 moderate, 56 high, 17 very high; Supplementary material, Table S13; Figure 6A). For the CLL EBV– patients, 36 significant correlations were found, with 7 negative (3 low, 4 moderate, 1 high) and 29 positive (7 low, 22 moderate; Supplementary material, Table S13; Figure 6B).
Receiver operating characteristic curve analysis for Epstein–Barr virus reactivation as a diagnostic marker
ROC curve analysis was performed to assess whether the frequency of specific T and B lymphocyte subpopulations expressing TLRs and their serum concentrations, could serve as predictive markers of EBV reactivation. The analysis indicated high specificity and sensitivity for TLR9+, TLR3+, CD8+TLR4+, CD19+TLR4+, CD4+TLR7+, and CD4+TLR8+ expression in the CVID patients. In the CLL patients, CD8+TLR9+, CD19+TLR9+, and CD4+TLR7+ showed similar diagnostic values. Positive expression of TLR9 on CD4+ T cells and CD19+ B cells was highlighted in both the CVID and CLL patients with EBV reactivation. ROC analysis of serum sTLR levels showed high sensitivity and specificity for sTLR3, sTLR7, sTLR8, and sTLR9 in the CVID EBV+ vs CVID EBV– patients, and for sTLR3 and sTLR9 in the CLL patients. The comparison of CVID EBV+ with CLL EBV+ patients indicated that sTLR3, sTLR9, and, to a lesser extent, sTLR8, may be useful markers (Supplementary material, Table S15).
Discussion
EBV occurs in most of the world’s population as a latent infection, and in patients with CLL, the virus often reactivates, as also evidenced in the many years of research conducted by the head of our research team.22,23 Studies have shown variable rates of EBV reactivation in leukemia, ranging from approximately 10% to 50% of cases.24 EBV reactivation in CLL is associated with unfavorable clinical outcomes, including more advanced disease, higher lymphocyte counts, and higher risk of disease progression,25 which was also observed in this study. From a clinical point of view, it is also associated with poorer overall survival and treatment resistance in patients with CLL. EBV reactivation can disrupt the normal immune response in CLL. This has been observed to lead to impaired immune surveillance, reduced T‑cell function, and altered cytokine profiles, which may contribute to disease progression and reduced antitumor immunity.25-30 EBV infection in leukemic cells can induce genetic and epigenetic changes, leading to aberrant cell signaling pathways and dysregulated gene expression.31,32 It has been shown to affect the expression of key CLL‑related genes, such as ZAP‑70 and TCL1 and to affect the B‑cell receptor signaling pathway.33,34 However, the literature to date has largely failed to mention the role of TLR‑based signaling pathways during EBV reactivation in the course of this SID. Our results showed that the immune response was impaired, involving almost all TLRs. Additionally, the observed changes in the percentage of selected T and B lymphocytes showing positive expression of the analyzed TLRs were significantly correlated in the patients with EBV reactivation. Individuals with CVID often have impaired immune function, particularly in terms of antibody production and cellular immune response to EBV. As a result, they have increased susceptibility to EBV infection and are at a greater risk of developing complications related to the virus.8,10,35 EBV can cause a variety of complications in patients with CVID, including chronic active EBV infection, EBV‑associated lymphoproliferative disorders, and malignancies, such as Hodgkin and non‑Hodgkin lymphoma. These complications can present with persistent fever, hepatosplenomegaly, lymphadenopathy, and other clinical symptoms.36-38 Dysregulated immune response to EBV can lead to continuous viral replication and dysregulated immune activation. This dysregulation includes altered cytokine profiles, impaired cytotoxic T‑cell function, and impaired natural killer cell activity, which contribute to the development of EBV‑related complications.39,40 Such immune dysregulation is observed in CVID, which may contribute to the inability of patients to mount an effective immune response against EBV. CVID patients often have deficiencies in the production of specific antibodies,41,42 including antibodies against EBV antigens, which was also shown in our study. The levels of specific antibodies against EBV in the patients with CVID were lower than in those with CLL. This defect additionally impairs their ability to control EBV infection and increases the risk of serious EBV‑related complications. Additionally, the deregulation of TLR‑based signaling pathways in the course of EBV reactivation in this specific group of PID patients has not been subject to previous. A few literature reports only indicate the participation of deregulation of selected TLRs in the course of CVID, but do not present mutual interactions or correlations with clinical aspects.42,43 In our study, we observed significant TLR pathway deregulation during EBV reactivation, aligning with previous findings indicating that EBV suppresses TLR9 expression via N6‑methyladenosine modifications. Zhang et al44 reported that the viral EBNA‑1 protein promotes methyltransferase‑like 3 degradation, a key methyltransferase for TLR9 mRNA stabilization, leading to reduced TLR9 expression and attenuated innate responses. This mechanism allows EBV to evade immune surveillance by targeting key innate immune receptors.
Future research should explore these interactions, particularly in immunodeficiency settings such as CVID and CLL, where TLR‑mediated immunity is already compromised. Increased serum TLR levels may have dual effects on immune responses. Higher TLR expression can enhance pathogen recognition, facilitating innate and adaptive immunity by promoting antigen‑presenting cell activation and T‑cell responses. However, excessive TLR activation may contribute to chronic inflammation, tissue damage, and diseases such as atherosclerosis and neurodegeneration. Additionally, overexpression of TLRs is linked to autoimmunity, where uncontrolled activation leads to immune responses against self‑tissues.
Prolonged TLR activation may also induce receptor desensitization, impairing the body’s ability to respond to new infections.44 Understanding these mechanisms is crucial for developing targeted therapies to modulate TLR pathways and improve immune homeostasis in patients with immunodeficiencies. Given the increased susceptibility to EBV‑related complications, regular screening for EBV infection and monitoring for symptoms of EBV‑related disease are essential in patients with CVID. In our study, EBV did not reactivate in the healthy individuals, as confirmed by serological tests and quantitative analyses, where viral copies remained undetectable. This indicates effective immune control, maintaining EBV in latency and preventing reactivation. Serological data suggest prior EBV exposure, highlighting the role of immune competence in suppressing reactivation. Differences in immune responses between healthy carriers and immunocompromised patients warrant further investigation. Regular monitoring, including imaging and serologic assessments, is crucial for early detection of complications. While EBV contributes to immunodeficiencies, not all infected individuals develop these conditions, suggesting the significance of genetic and other factors. Understanding EBV’s interaction with TLRs is essential for developing targeted therapies.
Our study has limitations. The sample size of the CVID and CLL patients was relatively small, affecting the generalizability of results. Patient heterogeneity in disease duration and severity may have influenced EBV reactivation and TLR dysfunction. Additionally, the cross‑sectional design prevented long‑term assessment of EBV dynamics. While we focused on TLR signaling, other immune pathways likely contribute to the observed dysfunctions. Finally, potential interactions with other infections were not evaluated, limiting a comprehensive interpretation of immune dysregulation in these patients. Future studies should address these gaps to refine therapeutic approaches.
Conclusions
In conclusion, this study provides important information on the impact of EBV reactivation on immune stability in patients with PIDs and SIDs. The results suggest that EBV reactivation is associated with dysregulation of TLR signaling pathways, which contributes to immune dysfunction and increased susceptibility to infections. Disruption of immune homeostasis has a significant impact on the quality of life of patients and the efficacy of therapy. The identification of this association highlights the need for further research on immunological mechanisms in patients with immunodeficiencies and the development of therapies aimed at restoring immune stability. This study may contribute to a better understanding of immunodeficiencies and the development of more effective treatment strategies and prevention of immunological complications.
- Abbott RJ, Pachnio A, Pedroza‑Pacheco I, et al. Asymptomatic primary infection with Epstein‑Barr virus: observations on young adult cases. J Virol. 2017; 91: e00382. | Crossref
- Chijioke O, Azzi T, Nadal D, Münz C. Innate immune responses against Epstein Barr virus infection. J Leukoc Biol. 2013; 94: 1185‑1190. | Crossref
- Merlo A, Turrini R, Dolcetti R, et al. The interplay between Epstein‑Barr virus and the immune system: a rationale for adoptive cell therapy of EBV‑related disorders. Haematologica. 2010; 95: 1769‑1777. | Crossref
- Callan MFC. The immune response to Epstein‑Barr virus. Microbes Infect. 2004; 6: 937‑945. | Crossref
- Houen G, Trier NH. Epstein‑Barr virus and systemic autoimmune diseases. Front Immunol. 2021; 11: 587380. | Crossref
SUPPLEMENTARY MATERIAL
ARTICLE INFORMATION